みんなの健康広場は生命の神秘を学ぶサイトです。
見出し西洋医学考察
骨格筋
骨格筋
|
骨格筋 (skeletal muscle) は、関節など骨格の可動部を動かす筋肉である[10]。脊椎動物では両端が腱を介して骨と繋がった形で配置され、昆虫やエビなどの節足動物ではクチクラ(角皮)を動かすために使われる。関節に関してその筋肉が収縮すると曲がるものを屈筋
、伸ばすものを伸筋
と言う。その他は回転筋・索引筋・括約筋などに分類される[10]。随意筋であるが、体躯の姿勢制御や反射などでは無意識に動く。体重比で成人男性の42%、同女性の36%を占める[11]。哺乳動物の骨格筋の密度は1.06kg/lであり、脂肪よりも約15%重い。 |
||||
関節関節(かんせつ)とは、骨と骨が連結する部分を表す言葉である。 脊椎動物の場合脊椎動物ではこれは内骨格における骨同士の連結部であり、不動性結合と可動性結合とがある。膝・肘・腰・肩など体中のいたるところにある。関節には、両骨の間に潤滑油の役目を果たす滑膜が存在する。
で結びつけられ、それらの牽引力によって曲がることができる。その動く方向、大きさはある程度決まっており、それを超える曲げ方があった場合には障害を起こす。ただし時間をかけて慣れさせれば可動範囲や方向をある程度変えることもできる。この幅は個体差も大きく、よく曲がる人は「関節が柔らかい」といわれる。これは、体操競技などでは重要な能力とされる。関節の可動幅が少ない人は怪我をしやすいともいわれる。 年齢を重ねるとリウマチなどで関節痛を感じることが多い。関節部分が損傷した場合、人工関節に置き換える治療法がなされる。 無脊椎動物の例棘皮動物は内骨格を持ち、ウミユリ類やウニ類などではそれらがつながって骨格系を構成するから、ここの骨格の間には一応関節があるが、骨格片が細かいため、脊椎動物のそれのような明瞭な関節とはなっていない。ただし、ウニ類の殻と針の間は、はっきりした関節になっており、これらは結合組織と筋肉で結びつき、根本から大きく動かすことができる。 節足動物では関節は外骨格のつなぎ目であり、それらの間は薄膜で繋がる。それはほぼ円周状になるが、対角線に当たる二点に両骨格間のジョイントがあり、関節面のそれを結ぶ線に直角方向に対して曲げることができる。 応用的な用法より広い意味では、その位置で折れるようになっている節のことを関節という。たとえばシダ植物では葉の基部に関節があれば、葉が枯れた際のその位置でぽろりと折れるが、無い場合には葉の基部が茎に張り付いて残る。これがあるか無いかはこの群では重要な分類上の特徴となっている。 関節の病気· 関節炎 · 変形性膝関節症 · 関節リウマチ · 痛風 · 偽痛風 · 顎関節症 · 感染性関節炎(化膿性関節炎) - 細菌感染が原因 |
平滑筋]
平滑筋 (smooth musle) は、横紋が無い筋肉であり、脊椎動物では心臓を除く内臓および血管を構成する筋肉である。無脊椎動物の身体を構成する筋肉はほとんどが平滑筋である[14]。収縮する速度は遅く数十秒かかる場合もあるが、一方で伸び縮みする率は大きく、その状態を保持する能力に優れる[14]。自律神経系から運動の促進・抑制双方の制御を受けている[14]。
心筋]
心筋 (cardiac muscle) は心臓を構成する筋肉である。心筋の特徴として、動作に必要な神経繊維が通常の神経繊維ではなく、特殊心筋と呼ばれる筋群によって興奮が伝達される。従って、肉眼的には神経繊維は存在しない。
|
洞房結節 刺激伝導系 > 洞房結節
心臓の内部図。洞結節の名称はないが、その領域はcrista terminalis(分界稜)付近に見ることができる。 洞房結節(どうぼうけっせつ、ラテン語: nodus sinuatrialis、英語: Sinoatrial node, 洞結節)は、心臓の右心房付近にあるペースメーカーの役目をする部分。発見者の名前からキース・フラック結節(Keith-Flack node)とも呼ばれる。70回/分のリズムを作る刺激伝導系の初端であり、生理的な心臓の収縮を指令する。その後刺激は房室結節へと伝わる。 房室結節 房室結節(ぼうしつけっせつ、田原結節、田原-アショフ結節、アショフ-田原結節、英 atrioventricular node: AV node)は、心臓における刺激伝導系の一部である。
赤線:刺激伝導系。房室結節は心房と心室の間にある。 田原淳がルードヴィッヒ・アショフの研究室で発見した。右心房の下方で心室中隔近くにある。洞房結節から房室結節までを刺激が伝わる速度(毎秒約1 m程度)や、プルキンエ線維などを刺激が伝わる速度(毎秒2 mを超える)と比べて、房室結節では毎秒約5 cmから、速くとも毎秒約10 cmと刺激伝導速度が10分の1以下と大変に遅い。また、刺激伝導の上で心房と心室は結合組織によって絶縁されており、心房の固有心筋の間を伝わる刺激は、ケント束のような奇形がある場合を除いて、心室の固有心筋へは直接伝わらないようになっている。したがって、房室結節へ伝わった刺激が、刺激伝導の上で心房と心室とを隔ている結合組織を唯一貫いているヒス束を伝わることで、心室側へと刺激が伝導される。以上の理由により、心房と心室の収縮に時間差を持たせることができる。正常な状態では心房からの興奮をヒス束に伝えるが、洞房結節の作るリズム(70回/分)が途絶えた場合、房室結節の持つ自動能が、調律の機能をある程度補完する。しかし、その時に作り出すリズムは40回/分程度と遅く、徐脈となる。 |


微細構造
筋肉の機能は、神経の制御を受けながら収縮する事と、その収縮度合いを測定しフィードバックすることである。ここでは主な構成要素を、骨格筋を例にして解説し、後に心筋と平滑筋の違いを述べる。
筋繊維(筋線維)
骨格筋を構成する細胞単位。筋芽細胞の融合によって生じる、細長く大きな巨大多核細胞
合胞体合胞体(ごうほうたい)またはシンシチウム(syncytium, pl. syncytia)は、動物に見られる、複数の核を含んだ細胞のこと。これに対して原生生物や菌類に見られる、一つの細胞に多数の核を持つ状態のものは多核体と呼ばれ、区別される。 合胞体は、数個から数千個もの核を含んだ細胞質の塊とも呼べる、一つの巨大な細胞である。合胞体が形成されるメカニズムは大きく二つに分けられる。一つは不完全な細胞分裂によって一個の細胞内に複数の核が作られる場合、もう一つは正常に形成された細胞同士が細胞融合を起こして複数の核を持つ巨大な細胞になる場合である。前者には昆虫の初期胚形成が、後者には骨格筋繊維の形成や哺乳類の胎盤、ウイルス感染細胞が、それぞれ代表的な例として挙げられる。なお、多核体は普通はこの前者の型に当たる。 合胞体の例昆虫の初期胚昆虫の多くでは不完全な細胞分裂が初期胚で行なわれ、細胞質分裂なくして核が大きな細胞質の中で複製される。 ショウジョウバエの様な無脊椎動物の初期胚シンシチウムはその細胞分化の特異化に重要である。卵細胞の細胞質はBicoidやNanosタンパク質をコードするmRNA分子の濃度勾配を持つ。Bicoidタンパク質は前端に集中し、Nanosタンパク質は後端に集中する。初期胚の核は迅速に分裂して、異なったBicoidとNanosの濃度を持った細胞質へ分散する。Bicoidの多い細胞質の核は胸部へ分化し、Nanosの多い細胞質の核は腹部へ分化する。 骨格筋の繊維形成大きな骨格筋繊維は数千の筋細胞が融合してできた合胞体である。骨格筋の合胞体は筋肉全体の迅速な調節された収縮を可能にする。運動ニューロンから伝えられた電気信号(活動電位)は、神経と筋が接合するシナプス(神経筋接合部)を介して筋繊維に伝えられる。電気信号は信号を受けた細胞膜の表面を素早く伝導するため、合胞体化して一つの細胞になった筋繊維は、それぞれがばらばらのときに比べてスムーズな伝達および運動が可能であると考えられている。 哺乳類の胎盤哺乳類の胎盤も代表的な合胞体の一つである。胎盤では、母体の血流に接する胚由来細胞が互いに融合して合胞体を形成する。この合胞体は、胚と母体の間の細胞の移動を制限するための障壁としての機能を果たしていると考えられている。血液細胞には上皮細胞同士の隙間を通過する事ができるように特化したものがあるが、胎盤上皮で形成された合胞体はそのような細胞の隙間(細胞間隙)を塞ぎ、母体の血流からの侵入を止める働きを担っていると考えられている。 ウイルス感染細胞の合胞体
ウイルス感染による合胞体 合胞体は、様々なウイルスが細胞に感染した時に、細胞同士が融合することで形成される場合がある。特にヒトヘルペスウイルスや、ヒト免疫不全ウイルス(HIV)、パラミクソウイルス(例 RSウイルス(RSV))が知られている。この細胞融合は、ウイルスが侵入した後、感染細胞の細胞膜表面に発現したウイルスの膜タンパク質の働きによって、隣接した非感染細胞との融合を起こすもの(fusion from within:感染が必須な細胞融合)と、ウイルス粒子が宿主細胞の細胞膜に結合して侵入しようとするとき、隣接する細胞同士を融合させるもの(fusion from without:ウイルス増殖を必要としない細胞融合)に大別される。合胞体を形成するかどうかは、そのウイルスと感染する細胞の種類によって決まり、ヒト免疫不全ウイルスなどのレトロウイルスや、ヘルペスウイルスでは前者のみ、センダイウイルスに代表されるパラミクソウイルスでは前者と後者の両方の現象が見られる。合胞体の形成は、これらのウイルスが細胞に感染したことを示す細胞の形態変化を示す特徴(細胞変性効果)の一つとして、ウイルス学の分野で利用されている。またセンダイウイルスの持つfusion from withoutは、さまざまな種類の動物細胞同士を融合させる手段として、バイオテクノロジーの分野で応用されている。 |
である。骨格筋が発生し分化する過程で、単核の筋原細胞同士が融合してつくられる。
筋原繊維(筋原線維
横紋筋の筋繊維中に存在する収縮性の構造体で、細胞内器官。直径約1μmの円筒状をしており、骨格筋では筋肉の長方向に沿って多くの筋原繊維が並行に並んでいる。微細な構造は、多くのサルコメアが厚さ2〜8nmのZ膜(Z線)と呼ばれる隔膜で仕切られながら10nm間隔で連結している。横紋筋の縞模様はこの並びが見えている[16]。ミオフィブリル、筋フィラメント、ミオフィラメントとも呼ばれる]。
サルコメア(筋節)
筋原繊維の最小構成単位。これが縦につながったものが筋原繊維である。個々のサルコメアは、ATP存在下で収縮が起こる。骨格筋の縞は、このサルコメアのアクチンフィラメントとミオシンフィラメントが並行に一部分が重なっている配列に由来する。筋小胞体から放出されたカルシウムイオンによりアクチンフィラメントがミオシンフィラメントな間に滑り込み筋肉が収縮する。したがって、そのときにはサルコメア全体の長さはアクチンフィラメントが滑り込んだ分だけ小さくなる。
サルコメアには、中央部に密度が高いA帯と、両側に密度が低いI帯がある。A帯は約1.5μm長のミオシンフィラメントで構成され、Z膜に接続したアクチンフィラメントがA帯に入り込んでいない部分がI帯である]。両フィラメントは、中心にあるミオシンフィラメントを六角形状にアクチンフィラメントが取り囲んだ断面構造を持つ。ミオシンフィラメント同士の中心間距離は40〜50nm、取り囲むアクチンフィラメントまでの距離は約15nmである]。
エネルギー
筋繊維はアデノシン三リン酸 (ATP) を使い、フィラメント同士がお互い重なり合うように引き付け合い収縮する[1]。
筋肉の制御
筋肉は、神経からの刺激で収縮を行っている。神経と筋肉は、神経筋接合部というシナプスの一種を介して刺激の伝達を行っている。神経末端からは、アセチルコリンが放出され、筋肉の側にあるアセチルコリン受容体に結合し、筋線維の細胞膜を脱分極させる。これがT管系を伝わって筋全体に広がり、T管系に接する筋小胞体
筋小胞体 図のSarcoplasmic
reticulumが筋小胞体 図のSarcoplasmic
reticulumが筋小胞体
筋小胞体(きんしょうほうたい、英: sarcoplasmic reticulum)は、筋線維内に存在する膜系構造の1つで、滑面小胞体のうち、収縮刺激伝達系として特殊化したものである。 筋小胞体は筋原線維と平行に存在する。弛緩した筋ではカルシウムイオンを含み、カルシウムイオンの放出によって筋を収縮させる。筋小胞体の間には横行小管が位置する。横行小管と接する部分は終末槽(英: terminal cisternae)と呼ばれる。 機能
である。 |
からカルシウムが放出される。このカルシウムをシグナルとして、アクチン繊維とミオシン繊維の間の滑り運動が起こるのである。
その他
筋繊維は本来積極的に伸展する能力は無く、弛緩したときに伸展するのは、骨格筋の場合、対立筋の働きによる外的な作用による。運動後の筋肉の疲労は、解糖系の最終生成物である乳酸によってもたらされるとの説があるが、医学的根拠は無い。
心筋の微細構造
心筋は、普通心筋と特殊心筋に分類される、特殊心筋としては、洞房結節、房室結節、ヒス束等が挙げられる。特殊心筋の働きは、心筋の統合された収縮を目的とした、興奮の伝達である。普通心筋は、骨格筋と同じように横紋があるが、骨格筋ほど整然と並んでは居ない。
平滑筋の微細構造
平滑筋を構成する細胞は紡錘形状で単一の核を持つ[14]。アクチンフィラメントを大量に持ち、ミオシンフィラメントは少量が不規則に分散している。細胞の形状はデスミン中間径フィラメントが存在して保たれる[14]。収縮にはカルシウムイオンによって制御されるが、小胞体があまり発達していないため、細胞膜にあるくびれの外側にイオンを溜め込んでいると考えられる[14]。
筋収縮や弛緩のメカニズム
大脳に発する運動指令は、小脳において修飾されたのち、遠心性の運動神経を介して、活動電位として伝えられ、運動神経と筋肉の連接部である神経筋接合部に至る。
運動神経の末端にある神経終末(シナプス前末端)に活動電位が伝わると、ここに分布する電位依存性Caチャネルを開口させて、Ca電流を生じる。これによるCa濃度上昇はACh(アセチルコリン)放出を惹起させ、ここで放出されたAChは、シナプス間隙に拡散して、筋肉側で神経終末と結合している終板に達する。終板にはAChのニコチン受容体があり、これにAChが結合することでNa(ナトリウム)、K(カリウム)、Ca(カルシウム)が流入して、いわゆる終板電位 (EPP)を発生させる。これは、筋鞘を介して筋線維全体に伝播されたのち、横行小管 (T管)を介して筋線維の中に入って筋小胞体へ至り、筋小胞体からCa2+の放出を引き起こす。これにより細胞内Ca2+濃度が増加し、トロポニンとCa2+が結合し、トロポニンにアロステリックな変化が生じる。この変化によりトロポミオシンが動き、ミオシンの作用部位が露出する。これによりミオシンとアクチンが反応して相対的な滑りを起こし、筋収縮が引き起こされる[1]。一方、Ca2+は、筋小胞体膜上のCa-ATPaseによって回収され、これによってCa濃度が正常値まで低下するとトロポニンとCa2+の結合が解除され、連鎖的に筋収縮は終了する。
なお、原生動物の組織内にもアクチンやミオシンがフィラメント状に存在している[1]。
筋タンパク質
脊髄動物の骨格筋には、湿潤重量で約20%のタンパク質が含まれ、これを筋タンパク質または筋肉タンパク質という。筋タンパク質の半分は細胞組織である細胞膜・ミトコンドリア・小胞体・細胞核などと、酵素タンパク質が占める。あとの半分は筋原繊維をつくる構造タンパク質であり、アクチン・ミオシンと調整タンパク質・骨格タンパク質などがある。
発生・発達
すべての筋肉は沿軸中胚葉から発生している。沿軸中胚葉は胎児の体躯に沿い、体節ごとに分かれている。これは主に3つがあり、脊髄を形成する硬節、皮膚を形成する皮膚分節、筋肉を形成する神経節である。この中で神経節は上下の節に分かれており、それぞれ軸上と軸下の筋肉へとなる。ヒトの場合、上分節は脊柱起立筋と椎間筋肉の一部にしかならない。手足を含むその他の筋肉は全て下分節から発達する]。
発生の期間、筋原繊維(筋前駆細胞)は脊椎に関連する筋肉へなるものと、その他の全筋肉を構成するため一度移動して体に取り込まれるものとに分かれる。通常では、側板中胚葉でつくられた筋原繊維がまず外郭を構成する結合組織を作る。そして筋原繊維は化学的な刺激に従いながら、それぞれ適切な場所で骨格筋を形成し始める。
生後、思春期前までは筋肉の発達に男女差が無いが、男性で思春期を迎えると第二次性徴によってアンドロゲンの分泌が活発となり、肩幅が広くなった後に筋肉が発達するようになり[18]、男女間に筋肉の差が生じるようになる。
筋肉は少なくとも2度大きな進化を遂げた。ひとつは刺胞動物であり、もうひとつは左右相称動物である。これらは海綿動物に相当するような有機体にある収縮可能な細胞から進化したものと考えられる[19]
数値
主な動物の筋肉繊維の直径
数値は David 1977 から。
|
動物 |
筋肉 |
直径(μm) |
|
赤筋繊維 |
36.0 |
|
|
(同) |
白筋繊維 |
49.4 |
|
脚筋 |
169.0 |
|
|
胸筋 |
26.8 |
|
|
横隔膜 |
18.0 |
|
|
ふくらはぎ(腓筋) |
60.8 |
|
|
長指伸筋 |
85.0 |
|
|
(同) |
横隔膜 |
34.0 |
|
横隔膜 |
25.0 |
|
|
横隔膜 |
30.0 |
|
|
横隔膜 |
60.0 |
|
|
横隔膜 |
34.0 |
|
|
(同) |
肋間筋 |
50.4 |
|
(同) |
三角筋 |
54.2 |
主な動物の筋力
数値は Prosesser 1973 から。
|
動物 |
筋肉 |
筋力(N/cm2) |
|
貝柱 |
117.7 |
|
|
指伸筋 |
29.4 |
|
|
遅下制筋 |
27.5 |
|
|
横隔膜 |
20.6 |
|
|
縫工筋 |
19.6 |
|
|
tenuissimus muscle |
13.7 |
|
|
気管平滑筋 |
7.8 |
筋肉の退化
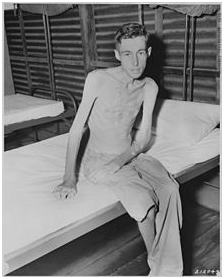
戦争捕虜。栄養失調の結果、筋肉に退化が見られる。その他にも、身体を活発に動かさないことや加齢、病気なども筋肉の退化の原因となる。
哺乳動物において、不活動や飢餓は骨格筋の退化に繋がる。筋肉量の減少は、タンパク質含有量の低下とともに筋肉細胞の減少や矮小化が伴う[22]。
ヒトでは、ベッドにずっと寝ていて運動を長期間行わない状態にあったり、宇宙飛行士が宇宙に滞在したりすると筋肉の弱化や退化が発生する事が知られている。有人宇宙飛行の世界では特に問題視され、無重力空間で過ごした結果、約30%もの筋肉が減ってしまったこともある[23][24]。これはヒトだけではなく、キンイロジリスやオオヒゲコウモリでも同様に起こると報告されている[25]。
年齢とともに、骨格筋の機能や量を維持する能力がゆっくりと減少する現象があり、これは筋肉減少症と呼ばれる。原因ははっきりしていないが、骨格筋繊維の維持を助ける「衛星細胞」が徐々に減ってゆく事との関連が疑われ、さらに筋肉量や衛星細胞を維持するために必要な感度もしくは分泌される重要な成長因子の減衰が関係すると考えられる。筋肉減少症は加齢によって一般的に起こり、また実際には病状として扱われないが、高齢者が怪我をしやすくなったり生活に支障を来たしたりする可能性がある]。